
Talk Overview
Meyerowitz begins his talk by reminding us of the critical role that plants play in human and global health. Undernutrition is a huge global problem that is only likely to worsen as the world’s population grows and global warming reduces arable land and plant yields. To breed for specific traits such as plant size, growth rate or yield, we urgently need to better understand plant biology; this includes the ability to predict the effects of genetic changes on plant traits. Meyerowitz and his collaborators have developed live imaging techniques to record dynamic changes in plant development, including changes in gene expression, protein localization, and phyllotaxis.
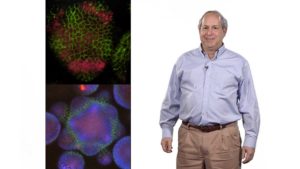
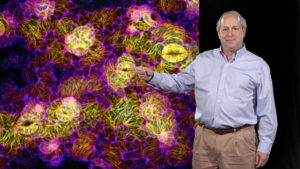
In Part 2, Meyerowitz describes how his lab is using live imaging to study phyllotaxis, the regular positioning of leaves or flowers around the developing apical shoot meristem. The plant hormone auxin has been known for years to induce flower and leaf growth. Meyerowitz and colleagues tracked the concentration of auxin and the auxin transporter protein, and the position of flower growth. Using this information they were able to develop a computer model that accurately reproduced phyllotaxis and auxin gradients in the apical meristem. The model also can predict how changing parameters such as rates of auxin diffusion or meristem size, will impact phyllotaxis and flower patterning.
The model described in Part 2 for determining phyllotaxis, requires that cells sense the concentration of auxin in neighboring cells. How do they do this? In his last lecture, Meyerowitz presents some fascinating data indicating that increased auxin concentration in a cell causes it to grow and expand, and expansion produces a physical force on neighboring cells. This mechanical force is sensed by cells and causes a redistribution of both microtubules and the auxin transporter forming a positive feedback loop. This is an amazing example of cell morphology influencing cell-cell communication, gene expression, and cell behavior.
Speaker Bio
Elliot Meyerowitz

Elliot Meyerowitz received his AB degree from Columbia and his PhD from Yale University. He was a post-doctoral fellow at Stanford and in 1980 he joined the faculty of The California Institute of Technology. Currently, Meyerowitz is the George W. Beadle Professor of Biology at Caltech and a Howard Hughes Medical Institute/ Gordon and Betty… Continue Reading




Leave a Reply